
 دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما هفته پنجم فروردین ماه 1403
دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما هفته پنجم فروردین ماه 1403 دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما اردیبهشت ماه 1403
دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما اردیبهشت ماه 1403 دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما هفته اول اردیبهشت ماه 1403
دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما هفته اول اردیبهشت ماه 1403 دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما هفته سوم اردیبهشت ماه 1403
دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما هفته سوم اردیبهشت ماه 1403 مجموعه 2 عددی طرح معرق خانه
مجموعه 2 عددی طرح معرق خانه برنامه اکسل متره و برآورد،تهیه صورت وضعیت راه و باند سال1403
برنامه اکسل متره و برآورد،تهیه صورت وضعیت راه و باند سال1403 دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما هفته دوم اردیبهشت ماه 1403
دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما هفته دوم اردیبهشت ماه 1403 سوالات نهاد مبحث دانش خانواده و جمعیت (بخش اول)
سوالات نهاد مبحث دانش خانواده و جمعیت (بخش اول) دانلود کتاب صوتی واپسین گفتار اسپالدینگ
دانلود کتاب صوتی واپسین گفتار اسپالدینگ پاورپوینت اثرات روانی خنده درمانی
پاورپوینت اثرات روانی خنده درمانی پاورپوینت کامل فصل سوم ریاضی چهارم ضرب و تقسیم(همراه با حل تمارین)
پاورپوینت کامل فصل سوم ریاضی چهارم ضرب و تقسیم(همراه با حل تمارین) دانلود رایگان کتاب صوتی از سکس تا فراآگاهی
دانلود رایگان کتاب صوتی از سکس تا فراآگاهی دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما هفته چهارم اردیبهشت ماه 1403
دانلود فایلهای بسته آمادهچاپ و نصب تابلو اعلانات مسجدنما هفته چهارم اردیبهشت ماه 1403 کیس اختلال دوقطبی - مصاحبه با بیمار دو قطبی - نمونه مصاحبه با بیمار دوقطبی
کیس اختلال دوقطبی - مصاحبه با بیمار دو قطبی - نمونه مصاحبه با بیمار دوقطبی پاورپوینت کیست هموراژیک تخمدان و درمان آن
پاورپوینت کیست هموراژیک تخمدان و درمان آن![دانلود پاورپوینت در مورد [سلول های بنیادی] - شامل 4 فایل مختلف - قابل ویرایش و ارائه - ppt](https://4kia.ir/s4/img_project/22638_1661011327.jpg) دانلود پاورپوینت در مورد [سلول های بنیادی] - شامل 4 فایل مختلف - قابل ویرایش و ارائه - ppt
دانلود پاورپوینت در مورد [سلول های بنیادی] - شامل 4 فایل مختلف - قابل ویرایش و ارائه - ppt پاورپوینت بازسازی ویرانه ها درس 22 مطالعات اجتماعی پنجم
پاورپوینت بازسازی ویرانه ها درس 22 مطالعات اجتماعی پنجم پاورپوینت بازی زندگی است درس 17 تفکر و سواد رسانه ای دهم
پاورپوینت بازی زندگی است درس 17 تفکر و سواد رسانه ای دهم مجموعه 4 عددی طرح معرق جنگنده
مجموعه 4 عددی طرح معرق جنگنده قالب آماده رنگی پلاک خودرو word
قالب آماده رنگی پلاک خودرو word![دانلود حل المسائل [طراحی و تحلیل آزمایش]: ویرایش هشتم - داگلاس مونتگومری ( 8 ) - زبان انگلیسی - pdf](https://4kia.ir/s4/img_project/22638_1659774456.jpg) دانلود حل المسائل [طراحی و تحلیل آزمایش]: ویرایش هشتم - داگلاس مونتگومری ( 8 ) - زبان انگلیسی - pdf
دانلود حل المسائل [طراحی و تحلیل آزمایش]: ویرایش هشتم - داگلاس مونتگومری ( 8 ) - زبان انگلیسی - pdf پاورپوینت درس هجدهم کارگاه کارآفرینی و تولید پایه دهم تعیین نوع کسب و کار
پاورپوینت درس هجدهم کارگاه کارآفرینی و تولید پایه دهم تعیین نوع کسب و کار پاورپوینت درس 2 علوم تجربی پایه چهارم دبستان (ابتدایی): مخلوط ها در زندگی
پاورپوینت درس 2 علوم تجربی پایه چهارم دبستان (ابتدایی): مخلوط ها در زندگی دانلود حل المسائل کتاب انتقال حرارت هدایتی دیوید هان و نجاتی اوزیشیک David Hahn
دانلود حل المسائل کتاب انتقال حرارت هدایتی دیوید هان و نجاتی اوزیشیک David Hahn طرح معرق کشتی هانسیاتیک
طرح معرق کشتی هانسیاتیک حل المسائل کتاب معادلات دیفرانسیل و مسائل مقدار مرزی دیپریما و بویس ویرایش نهم William Boyce
حل المسائل کتاب معادلات دیفرانسیل و مسائل مقدار مرزی دیپریما و بویس ویرایش نهم William Boyce پاورپوینت کامل و جامع با عنوان تئوری رفتار مصرف کننده در اقتصاد در 112 اسلاید
پاورپوینت کامل و جامع با عنوان تئوری رفتار مصرف کننده در اقتصاد در 112 اسلاید پاورپوینت پودمان هفتم کار و فناوری پایه هفتم کار با چوب
پاورپوینت پودمان هفتم کار و فناوری پایه هفتم کار با چوب دانلود جزوه الاستیسیته اصغری شریف
دانلود جزوه الاستیسیته اصغری شریف فروش فیلتر بورسی استریکلی فقط 75 هزار تومان
فروش فیلتر بورسی استریکلی فقط 75 هزار تومان کسب درآمد اینترنتی 300000 تومان در خانه در کمتر از 30 دقیقه
کسب درآمد اینترنتی 300000 تومان در خانه در کمتر از 30 دقیقه روش درآمدزایی در خواب (تعجب نکنید! کلیک کنید و بخوانید)
روش درآمدزایی در خواب (تعجب نکنید! کلیک کنید و بخوانید) ربات همه کاره اینستاگرام
ربات همه کاره اینستاگرام کسب و کار اینترنتی با درآمد میلیونی
کسب و کار اینترنتی با درآمد میلیونی كسب درآمد اينترنتي روزانه حداقل100هزار تومان تضميني
كسب درآمد اينترنتي روزانه حداقل100هزار تومان تضميني کسب درآمد ابدی و بی نهایت 100% واقعی
کسب درآمد ابدی و بی نهایت 100% واقعی کسب درآمد روزانه حداقل یک میلیون تومان ! کاملا حلال و واقعـی !!
کسب درآمد روزانه حداقل یک میلیون تومان ! کاملا حلال و واقعـی !! مجموعه ی آموزش تعمیر لامپ کم مصرف (از مبتدی تا پیشرفته)
مجموعه ی آموزش تعمیر لامپ کم مصرف (از مبتدی تا پیشرفته) افزایش ممبر کانال، گروه و ربات تلگرام به صورت بی نهایت (اد ممبر)
افزایش ممبر کانال، گروه و ربات تلگرام به صورت بی نهایت (اد ممبر) آموزش بازكردن انواع قفل ها بدون كليد(ويژه)
آموزش بازكردن انواع قفل ها بدون كليد(ويژه) کسب و کار اینترنتی در منزل
کسب و کار اینترنتی در منزل آموزش برنامه نویسی آردوینو
آموزش برنامه نویسی آردوینو دانلود مجموعه آموزشی پایپینگ ( Piping ) و نقشه خوانی + آموزش سه نرم افزار طراحی و تحلیل لوله کشی صنعتی
دانلود مجموعه آموزشی پایپینگ ( Piping ) و نقشه خوانی + آموزش سه نرم افزار طراحی و تحلیل لوله کشی صنعتی بازگردانی پیامک های حذف شده- ریکاوری پیامک ۱۰۰٪ عملی
بازگردانی پیامک های حذف شده- ریکاوری پیامک ۱۰۰٪ عملی آموزش رایگان کسب درآمد از سایت الیمپ ترید ( olymp trade )
آموزش رایگان کسب درآمد از سایت الیمپ ترید ( olymp trade ) اموزش ویرایش امضا و پکیج برنامه اندروید و کسب درامد از مارکت های اندرویدی
اموزش ویرایش امضا و پکیج برنامه اندروید و کسب درامد از مارکت های اندرویدی کد های آماده html و css جهت یادگیری و طراحی سریع
کد های آماده html و css جهت یادگیری و طراحی سریع دانلود نمونه فاکتور آماده با فرمت ورد - اکسل و عکس
دانلود نمونه فاکتور آماده با فرمت ورد - اکسل و عکس آموزش ساخت بازی بدون دانش برنامه نویسی و طراحی سه بعدی مبتدی تا پیشرفته با نرم افزار
آموزش ساخت بازی بدون دانش برنامه نویسی و طراحی سه بعدی مبتدی تا پیشرفته با نرم افزار اموزش کسب درامد از اینترنت روزانه ۳میلیون تومان تضمینی و تست شده
اموزش کسب درامد از اینترنت روزانه ۳میلیون تومان تضمینی و تست شده آموزش كامل تعمير لامپ كم مصرف(اختصاصي)
آموزش كامل تعمير لامپ كم مصرف(اختصاصي) نسخه خطی اشعار و پیشگویی های شاه نعمت الله ولی
نسخه خطی اشعار و پیشگویی های شاه نعمت الله ولی نسخه خطی اشعار و پیشگویی های شاه نعمت الله ولی
نسخه خطی اشعار و پیشگویی های شاه نعمت الله ولی درامدزایی در خواب! (تعجب نکنید! بخوانید)
درامدزایی در خواب! (تعجب نکنید! بخوانید)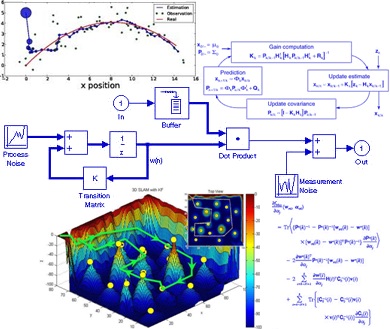 دانلود پاورپوینت فیلتر کالمن بر روی یک سنسور شتاب سنج برای تخمین سه متغیر حالت
دانلود پاورپوینت فیلتر کالمن بر روی یک سنسور شتاب سنج برای تخمین سه متغیر حالت مدار داخلی واکی تاکی(اموزش ساخت)
مدار داخلی واکی تاکی(اموزش ساخت) کتاب افزایش ممبر کانال تلگرام
کتاب افزایش ممبر کانال تلگرام اندیکاتور ای کیو آپشن
اندیکاتور ای کیو آپشنپیوند ها

دانلود مقاله ترجمه شده با عنوان "اثرات تنش شوري بروي اسيد آمينه، اسید آلی و ترکیب کربوهیدرات ریشه ها در گياه يونجه" - رشته کشاورزی - word
دانلود مقاله ترجمه شده با عنوان "اثرات تنش شوری بر روی اسید آمینه، اسید آلی و ترکیب کربوهیدرات ریشه ها، باکتروئید ها و سیتوزول یونجه (Medicago sativa L.)" برای رشته کشاورزی شامل 6 صفحه متن انگلیسی با فرمت pdf. دانلود از لینک زیر : 14 صفحه متن ترجمه فارسی با فرمت word
چکیده: اسید آلی محلول اتانول، کربوهیدرات و ترکیبات اسید آمینه ریشه ها و گره های (سیتوزول و باکتروئید) یونجه (Medicago sativa) توسط کروماتوگرافی گاز مایع و کروماتوگرافی مایع با عملکرد بالا شناسایی شده است. در میان اسیدهای آلی سیترات ترکیب غالب در ریشه ها و سیتوزول همراه با وجود مولانت در غلظت بالا در باکتروئید بود. این دو اسیدهای آلی همراه با مالات و سوکسینات برای بیش از 85 درصد مخزن اسید آلی در گره ها و برای 97 درصد در ریشه ها اختصاص یافتند. کربوهیدرات های اصلی در ریشه ها، سیتوزول گره و باکتروئیدها (ترتیب نزولی غلظت) بودند: ساکارز، پینیتول، گلوکز و اونونیتول. مالتوز و ترهالوز به نطر می رسد در غلظت های بسیار کمی موجود باشند. آسپاراژین، گلوتامات، آلاتین، y-aminobutyrate و پرولین اسید آمینه های اصلی در سیتوزول و باکتروئیدها بودند. علاوه بر این محلول ها، سرین و گلوتامین در ریشه ها به خوبی حضور داشتند و نشان داده شد. هنگامی که گیاهان یونجه به مدت 2 هفته در معرض تنش کلرید سدیم قرار گرفتند، تمامی غلظت اسید آلی در گره ها و ریشه ها تا بیش از 40 درصد افت کردند، در حالی که غلظت لاکتات به ترتیب در سیتوزول، ریشه ها و باکتروئیدها تا 11، 27 و 94 درصد افزایش داشتند. در باکتروئیدها، لاکتات به فراوان ترین اسید آمینه آلی تبدیل شد و ممکن است تا حدی به تنظیم اسمزی کمک کند. از طرف دیگر، تنش شوری باعث القای افزایش زیادی در اسید آمینه و مخزن های کربوهیدرات شد. در درون اسید آمینه ها پرولین بیشترین افزایش را نشان داد به ترتیب 3/11، 8/12 و 8 برابر در ریشه ها ، سیتوزول و باکتروئیدها. تجمع آن مکانیسم تنظیم اسمزی را نه تنها در ریشه ها بلکه در بافت گره منعکس می کند. به موازات آن، غلظت آسپاراژین تا حد زیادی افزایش یافت؛ این آمید املاح نیتروژن اصلی را باقی گذاشت و در باکتروئیدها نقش مهمی در تنظیم اسمزی ایفا کرده است. در مقابل، تیمار نمک اثر خیلی محدودی بر روی غلظت سایر اسید آمینه ها داشت. در میان کربوهیدرات ها، غلظت پینیتول به طور قابل توجهی به ویژه در سیتوزول و باکتروئیدها ( به ترتیب 4/5 و 4/3 برابر ) افزایش یافته بود، که این سیکلیتول برای بیش از 35 درصد از کل مخزن کربوهیدرات اختصاص یافته است. پینیتول ممکن است به مقاومت در برابر تنش شوری کمک کند. با این حال، غلظت ترهالوز هم در گره ها و هم در ریشه ها در سطح پایینی باقی می مانند و نقش آن در تنظیم اسمزی در یونجه به نظر بعید است. اثرات خاک های شور بر روی رشد گیاهان موضوع پژوهش های بسیاری در قریب به 100 سال بوده است به این دلیل که تنش شوری بهره وری محصول را با محدودیت مواجه می سازد. مقاومت به شوری گیاهان پدیده ی پیچیده ای است که شامل تغییرات مورفولوژیکی و توسعه ای و تکاملی و همچنین فرایندهای فیزیولوژیکی و بیوشیمیایی می شود. این جنبه ها به طور دوره ای در چندین بررسی مورد پوشش قرار گرفته است. بقا و رشد در محیط های شور نتیجه ی فرایندهای سازگاری نظیر انتقال یون و جداسازی آن ، سنتز و تجمع املاح اسمزی است که منجر به تعدیل اسمزی و گردش پروتئین برای احیای مجدد سلولی می شود. اگرچه این مکانیسم های آسیب شوری و مقاومت به شوری کل گیاه به طور گسترده ای مورد مطالعه قرار گرفته است لذا مطالعات در زمینه ی سطوح سلولی، اندامی و مولکولی هنوز آنچنان که باید و شاید صورت نگرفته است.
جهت کپی مطلب از ctrl+A استفاده نمایید نماید |
برچسب های مهم

بررسی اثر تنش خشکی روی بیان برخی از ژنهای آنتیاکسیدانت درگیاه دارویی کنجد sesamum indicum L.) (
چکیده خشکی به عنوان يکی ازمهم ترين عوامل محدود کننده رشد و توليد گياهان زراعی شناخته شده است. تغييرات فيزيولوژيکی که در اثر کمبود آب در گياه ايجاد میشود و صدمات اکسيداتيو بيشترين صدمات را به گیاه وارد می کنند. آنزيمهاىآنتىاكسيدانازجملهمهمترينراهكارهاىدفاعىگياهاندرمهارگونههاىاكسيژنفعالمىباشند. دراينمطالعه،بهجهت بررسی اثر تنش خشکی روی بیان ژن های آنتی اکسیدانت و میزان فعالیت آنزیم های آنتی اکسیدانت در گیاه دارویی کنجد انجام گرفت. ژن های مورد بررسی ، ژن های کاتالاز ، آسکوربات پراکسیداز و پلی فنیل اکسیداز بودند . ژن های بتا اکتین و آلفا توبولین نیز به عنوان ژن های رفرنس انتخاب شدند. به منظور بررسی اثر تنش در 6 دوره زمانی نمونه برداری صورت گرفت. پس از استخراج RNAو ساخت cDNA بررسی بیان ژن با استفاده از QRT-PCR انجام شد. پسازآنپاسخآنزيمى،بياننسبىژنهاىكاتالاز،آسکوربات پراکسیداز و پراکسیداز وبرخىازشاخصهاىفيزيولوژيكى سنجيدهشد. میزان بیان ژن آسکوربات پراکسیداز با گذشت زمان افزايش یافته در مقابل میزان بیان کاتالاز تغییرات منظمی مشاهده نشده است .بيشترفعاليتآنزيمهاىآنتىاكسيداندركناربيانپايدارتروبيشترژنهاى مورد نظرمشاهده شد. افزايشميزانبيانژنآسكورباتپراكسيدازنشاندادكهاينآنزيممىتوانددربرابرتنشخشکیپاسخنشاندادهوكمبودكاتالازراجبراننمايد. نتایج حاصل نشان داد که مقاوم ترین آنزیم برای مقابله با تنش آنزیم آسکوربات پراکسیداز می باشد. این آنزیم از همان روزهای ابتدای تنش افزایش فعالیت نشان داده ولی با افزایش زمان به خاطر آسیب به ساختارهای تولید کننده ی آنزیم ها، کاهش معنی داری را پیدا می کند. افزایش تحمل به خشکی با افزایش بیان ژن های کد کننده ی آنزیم های آنتی اکسیدان همراه شده است. مطالعات نشان می دهد همبستگی بین تحمل به خشکی و ظرفیت دفاع آنتی اکسیدانت وجود دارد. فهرست مطالب
شکل2-1 ....................................................................................................................................54 شکل 3-1...................................................................................................................................59 شکل 4-1 ...................................................................................................................................60 شکل 4-2....................................................................................................................................60 شکل 4-3..................................................................................................................................63 شکل 4-4...................................................................................................................................64 شکل 4-5...................................................................................................................................66 شکل 4-6...................................................................................................................................66 شکل 4-7..................................................................................................................................67 شکل 4-8..................................................................................................................................68 شکل 4-9..................................................................................................................................69 شکل 4-10..................................................................................................................................70 شکل 4-11..................................................................................................................................71 شکل 4-12 ................................................................................................................................72 شکل 4-13.................................................................................................................................72 شکل 4-14.................................................................................................................................73 فهرست جداول جدول2-1...................................................................................................................................46 جدول 3-1.................................................................................................................................57 جدول 3-2.................................................................................................................................57 جدول 3-3................................................................................................................................58 جدول 3-4.................................................................................................................................59 جدول 4-1...................................................................................................................................61 جدول 4-2 .................................................................................................................................61 فصل اول مقدمه 1-1 مقدمه کنجد (Sesamum indicum L.) گیاه یکساله، خودگشن با دیرینه زراعی طولاني که به علت دارا بودن درصد زیاد روغن و مقدار مناسب پروتئین بهعنوان یک منبع تغذیهای محسوب میشود (نصر، 1382). سطح زیر کشت جهاني كنجد در سال 2010 میلادی، در میان 10 کشور عمده تولیدکننده کنجد در جهان، کشور میانمار با تولید 000/720 تن در رتبه اول تولید قرار دارد. کشور هند با تولید 000/620 تن در رتبه دوم و چین با تولید 000/590 تن در رده سوم قرار دارد که در این بین ایران با تولید 31848 تن در رده دهم قرار دارد. متوسط عملکرد کنجد در جهان در این سال 490 کیلوگرم در هکتار بوده است (فائو، 2010). مناطق اصلی کشت کنجد در کشور استانهای فارس، خوزستان، جیرفت،بوشهر،اردبیل (مغان)،خراسان شمالی میباشد. پنج استان فارس، خوزستان، جیرفت،خراسان رضوی و اردبیل همهساله 80 درصد سطح زیر کشت کنجد کشور را به خود اختصاص دادهاند. در سال 1391 سطح زیر کشت کنجد در کشور به 43475 هکتار با تولید 39614 تن و عملکرد 912 کیلوگرم در هکتار بوده است. کنجد به اسامی Till, Simsim, Gingelly, Benni,Seed معروف است. كنجد از دانههای روغني مناطق گرم و نيمهگرم است، ولي كشت ارقام جديد آن به مناطق معتدله نيز گسترشیافته است. اين گياه داراي ارقام محلي زيادي است و در اغلب كشورها توسط كشاورزان خردهمالک و بهصورت سنتي كشت و كار میشود (وایز، 2000). مقدار روغن در زمان رسيدگي فيزيولوژيكي به سطح ثابتي میرسد و تا زمان رسيدگي بذر نوسان اندكي دارد (فلوئر و داونی، 1970). آبياري میتواند موجب افزايش مقدار روغن شود، درصورتیکه تنش خشكي (میلر و رایتن، 1987) موجب كاهش آن میشود و بهطورکلی عوامل محيطي اثرات معكوس بر مقدار روغن و پروتئين دارند. ازنظر درصد روغن و عملكرد روغن دانه ارقام مقاوم به تنش موفقتر عمل مینمایند (اسمیت، 1990). کشور ایرانازنظر اقلیمی در منطقه خشک نیمهخشک دنیا قرار دارد، ازاینرو خشکی یکی از مشکلات پیش روی زراعت کشور ماست. خشکی سبب تغییرات مورفولوژیکی، فیزیولوژیکی و بیوشیمیایی متعددی در گیاهان میشود ضمن اینکه تحمل به خشکی در گیاهان نیز خصوصیتی ثابت نبوده و ممکن است در مراحل مختلف رشد هرگونه، متفاوت باشد (سایرام،2001). بهطورکلی گیاهان، طیف وسیعی از تنشهای محیطی را که نهایت منجر به بروز تنش اکسیداتیو میشود، درک میکنند. مکانیسم مقاومت در برخی از تنشها بهصورت یک ارتباط درونی و نتیجه یک برنامهریزی هماهنگ و پیچیده است. در شرایط تنش عدم توازن بین فرآیند جذب انرژی و مصرف آن توسط اندام فتوسنتزی باعث تولید انواع اکسیژن فعال (ROS) و ناتوانی گیاه در مهار آن میگردد که درنهایت منجر به بروز تنش در غشای سلول و بروز علائم ناشی از صدمات اکسیداتیو شود (بلوخینا و همکاران، 2003). افزايش ميزان رادیکالهای فعال اکسيژن در گياه باعث ميشود که برای کاهش اثرات سمي تنش اکسيداتيو ناشي از تنش خشکی، مکانيسمهاي متنوعي در گياه فعال شود. در اين شرايط ميزان آنتیاکسیدانتها افزایشیافته و آنزیمهای مهارکننده ROSها در جهت کاهش اثرات سمي ناشي از تنش اکسيداتيو حاصل از تنش خشکی، افزايش پيدا ميکنند (كافي و همكاران،1382). برای مقابله با تنش اکسیداتیو آنزیمهای آنتیاکسیدانت نظیر کاتالاز (CAT)، پراکسیداز (POD)، آسکوربات پراکسیداز (APX)، سوپراکسید دیسموتاز (SOD) و گلوتاتیون ردوکتاز (GR) باعث حذف و غیرفعال شدن گونه های فعال اکسیژن میشوند (دیرینک و مونتاگو،2002). لذا بررسی اثرات تنش خشکی و الگوی رفتاری آنزیمهای آنتیاکسیدانت، میتواند به درک مکانیسمهای دخیل در القاء مقاومت یا تحمل به گیاه کمک نماید، چراکه همبستگی بالایی در تحمل به تنشهای محیطی و تغییرات غلظت آنزیمهای آنتیاکسیدانت در گیاهان فتوسنتز کننده وجود دارد. از سوی دیگر با عنایت به اینکه سنتز هر مادهای در سلول تحت کنترل ژنهای مسئول سنتز آن است انتقال این ژنها به گیاهان دیگر و تولید ارقام محتمل به خشکی دور از ذهن نخواهد بود. اهداف تحقیق حاضر عبارتاند از: 1- مقایسه الگوی بیان برخی ژنهای دخیل در تنش خشکی در سطح آنزیمشامل کاتالاز، سوپراکسید دیسموتاز و آسکوربات پراکسیدازو پراکسیداز در بافت برگ و ریشه 2- بررسی الگوی بیان برخی ژنهای دخیل در تنش خشکی در سطح ترانسکریپتوم 1-2 کلیات 1-2-1 گیاهان و تنشهای محیطی گیاهان در دوره حیاتشان با انواع تنشهای محیطی مواجه میشوند، این تنشها شانس نمو و بقای گیاهان را محدود میکنند. در بسیاری از نقاط کره خاکی شرایط مناسب رشد فقط برای مدت کوتاهی دوام دارد و گیاهان مجبورند که در همین زمان کم، مراحل اساسی رشد خود را انجام دهند در برخی نقاط هم که شرایط برای رشد مناسب است، افزایش تراکم و تعداد گیاهان عامل ایجاد رقابت برای گیاهان در به دست آوردن مواد غذایی، آب و نور است (لارچر و همکاران، 2001). درمجموع تنش یعنی شرایط نامناسبی که حتماً مرگ آنی در پی نداشته و بهطور دائم یا موقت در یک محل اتفاق میافتد ولی بر عملکردهای حیاتی موجودات تاثیرداشته باشد (والتر و همکاران، 1985). از قوانین حرکت نیوتن چنین استنباط شده است که اگر هر موجودی تحت تاثیرعملی (تنش) قرارگیری عکسالعملی (واکنش) از خود نشان میدهد. واکنش میتواند برگشتپذیر یا برگشتناپذیر باشد. اگر واکنش از شدت کافی برخوردار باشد موجود زنده دچار یک تغییر پایدار یعنی صدمه یا مرگ میشود. بههرحال دانشمندان علوم گیاهی تنش را با دو تعریف بومشناختی و بیوشیمیایی موردتوجه قرار میدهند. تنش در مفهوم بومشناختی: فشارهای زیستمحیطی است که نسبت تولید ماده خشک را در قسمتی از گیاه یا تمامی آن محدود میکند اما تنش ازنظر بیوشیمیایی به معنی اختلال در تولید طبیعی ترکیبات مختلف گیاهی است. امروزه تنش را به دو گروه طبقهبندی میکنند اول تنشهای زیستی[1]: تنشهایی که حاصل حمله یک موجود زنده به موجود زنده دیگر است مانند آفتها، پاتوژنها و آللوپاتی؛ دوم تنشهای غیر زیستی[2] شامل: 1- باد، فشار، صدا، نیروهای مغناطیسی و الکتریکی 2- شیمیایی، مثل شوری، یونی، علفکشها و... 3- تشعشع، مثل پرتوهای A-B, UV- UV 4-آب، مثل غرقابی و خشکی 5- دما، مثل گرماو سرما هر نوع از تنشها در وهله اول تنش اولیه محسوب شده و منجر به تغییراتی در سیستم زیستی میشود. اگر مدتزمان تنش اولیه کوتاه باشد اثرات آن در حد چند ثانیه یا دقیقه مشاهده میشوند؛ اما اگر مدتزمان بروز تنش طولانی باشد تنش ثانویهپدید آمده و آسیب حاصل از آن غیرمستقیم خواهد بود. گیاهان نیزمانندجانوران برای مقابله با این شرایط ناسازگارو سخت با استفاده از مکانیسمهای متفاوت با تنش مقابله مینمایند که این مکانیزمها شامل سازش و مقاومت بوده که مقاومت خود شامل تحمل کردن، اجتناب و فرار میباشد (پارساد، 1996). همه این تنشها، پاسخهای مشترکی را در گیاهان القا میکنند ولی در شروع سیگنال باهم تفاوت داشته و همچنین گیرندههای آنها متفاوت میباشند. این تنشها با بیان یک سری از ژنها گیاه راقادربه ترمیم آسیب ناشی از تنش کرده یا گیاه رادر برابر تنشهای محیطی که آینده پیش خواهد آمد حمایت میکنند (باجوز و همکاران، 2008)؛ یعنی درواقع گیاه را با محیط سازگار کرده و بقای آن را بیشتر تأمین و تضمین میکنند و از این طریق بهنوعی با محیط خود سازش یافته یا تطابق مییابند. سازشها اعم از مورفولوژیکی، فیزیولوژیکی و بیوشیمیایی به نحوی انجام میگیرد که گیاه بتواند بهتر با محیط تطابق پیدا کند و از امکانات مادی محیط خود به گونه شایستهای استفاده کند. بهطور مثال سازش بیوشیمیایی در گیاهان شامل انواع تغییرات بیوشیمیایی در سلول است. این تغییرات شامل تکامل راههای متابولیکی جدید، تجمع متابولیکهای با وزن مولکول کم، سنتز پروتئینهای خاص، مکانیسمهای سمزدایی و تغییر در میزان فیتوهورمونها میباشند (باجوز و همکاران، 2008). در اغلب تنشهای محیطی، تغییرات ایجادشده گیاهان را در مواجهه با تنش کمک میکنند. یکی از عمدهترین تنشهای محیطی که اغلب گیاهان با آن مواجهاند تنش خشکی میباشد. 1-2-2 تنش خشکی تنش خشكی زمانی در گیاه حادث میشود كه میزان آب دریافتی گیاه كمتر از تلفات آن باشد. این امر ممكن است به علت اتلاف بیشازحد آب یا كاهش جذب ویا وجود هر دو مورد باشد (کوچکی،1374). یكی از نشانههای كمبود آب، كاهش تورژسانس و درنتیجه رشد و توسعه سلول بهویژه در ساقه و برگها است. به همین دلیل اولین اثر محسوس کمآبی را میتوان از اندازه کوچکتر برگها یا ارتفاع گیاهان تشخیص داد. به دنبال كاهش سطح برگ، جذب نور نیز كم شده و باعث كاهش ظرفیت كل فتوسنتزی گیاه شده و رشد و درنهایت عملكرد آن دچار نقصان میشود (حسنی، 1385). خشكی بر جنبههای مختلف رشد گیاه تاثیر گذاشته و موجب كاهش و به تأخیر انداختن جوانهزنی، كاهش رشد اندامهای هوایی کاهش تولید ماده خشك میگردد. درصورتیکه شدت تنش خشکی زیاد باشد، موجب كاهش شدید فتوسنتز، مختل شدن فرآیندهای فیزیولوژیكی، توقف رشد و سرانجام مرگ گیاه میشود (سینگ و همکاران، 1996). كمبود آب ازمهمترین مشكلات مناطق خشك و نیمهخشک میباشد كه بر روی رشد و نمو گیاهان اثر میگذارد (اهدایی، 1372). نواحی خشك و نیمهخشک مناطقی هستند كه كل تعرق گیاهان در آن 50% تعرق در شرایط بدون تنش ویا حتی كمتر از این مقدار باشد. متأسفانه كمبود آب تنها منحصر به این نواحی نمیشود، بلكه حتی در شرایط آبوهوای مرطوب و یا توزیع نامنظم بارندگی نیز منجر به محدود شدن آب قابلدسترس و درنتیجه كاهش رشد گیاه میشود (کافی، 1384). در كشور ما بهجز سواحل دریای خزر و قسمتهای كوچكی از شمال غربی كشور بقیه مناطق جزء مناطق خشك و نیمهخشک محسوب میشوند. این در حالی است كه مناطق خشك كشور نسبت به مناطق نیمهخشک آن از وسعت بیشتری برخوردار است (اهدایی، 1372). 1-2-3 لزوم معرفی گیاهان متحمل به خشکی 5/۹۷درصد آبکره زمین در دریاها و دریاچهها است که آبشور را تشکیل میدهد. 5/۲ درصد باقیمانده آب شیرینی است که در زمین وجود دارد که از آن ۳/۰درصد آب رودخانهها ۸/۳۰ درصد آبهای زیرزمینی و ۹/۶۸ درصد یخچالها و پوشش دائمی برف کوهها است. کشور ايران به خاطر قرار گرفتن در كمربند خشك جغرافيايي و نوار بياباني كه در 25تا 40درجه عرض شمالي واقعشده است، از شرايط آبوهوایی برخوردار است كه جزو مناطق كم باران جهان بشمار میآید. باوجود قرارگرفتن کشور ايران در کمربند خشک جهانيتغییرپذیری شديد اقليمي، بارش تنها معادل یکسوم متوسط جهاني میباشد (حيدري شریفآباد و همکاران،1381). بر اساس گزارشها در سال 1380حدود 2/6ميليون هکتار زراعت آبي و 4ميليون هکتار زراعت ديم و 1/1ميليون هکتار از باغات تحت تاثیرداشته خشکسالی قرارگرفتهاند. خسارت ناشي از خشکسالی بر باغات در اين سال بالغبر 520ميليون دلار بود. بر اساس تحقيقات انجامگرفته در کشور، اثر مستقيم خسارت ناشي از کاهش هر 1میلیمتر بارندگي برابر 98ميليارد ريال است (غفاری،1386).ﺗﻨﺶ ﺧﺸﻜﻲ ﺑﻴﺶ از ﻫﺮ ﻋﺎﻣﻞ دﻳﮕﺮي ﻣﻮﺟﺐ ﻛﺎﻫﺶ رﺷﺪ و ﺗﻮﻟﻴﺪ ﮔﻴﺎﻫﺎن میگردد. ﺑﻬﺘﺮﻳﻦ راه ﻣﻘﺎﺑﻠﻪ ﺑﺎ ﺧﺸﻜﻲ اﺳﺘﻔﺎده ﺑﻬﻴﻨﻪ از آب و اﺻﻼح ﮔﻴﺎﻫﺎن ﺑﺮاي اﻓﺰاﻳﺶ ﻣﻘﺎوﻣﺖ ﺑﻪ ﺧﺸﻜﻲ اﺳﺖ. ﻣﻘﺎوﻣﺖ ﺑﻪ ﺧﺸﻜﻲ، ﺗﻮاﻧﺎﻳﻲ ﮔﻴﺎه ﺑﺮاي رﺷﺪ ﻣﻄﻠﻮب در ﺷﺮاﻳﻂ ﺗﻨﺶ است. 1-2-4 کاهش رشد گیاه تحت تنش خشکی ﺑﻴﺶ از 80 درﺻﺪ ﺑﺎﻓﺖ ﮔﻴﺎﻫﻲ را آب ﺗﺸﻜﻴﻞ میدهد (مارشال، 1986)و ﻛﻤﺒﻮد آن در ﮔﻴﺎﻫﺎن ﻋﻮارض ﺷﺪﻳﺪي را بهسرعت آﺷﻜﺎر میسازد ومهمترین ﻋﺎﻣﻞ ﻣﺤﺪودﻛﻨﻨﺪه رﺷد و ﻧﻤﻮ ﮔﻴﺎﻫﺎن ﻣﺤﺴﻮب میشود. ﺑﺮاي اﻳﻨﻜـﻪ ﮔﻴـﺎه ﺑﺘﻮاﻧـﺪ آب ﺟﺬب ﻛﻨﺪ، ﺑﺎﻳﺴﺘﻲ ﭘﺘﺎﻧﺴﻴﻞ آب سلولهای رﻳﺸﻪ ﻧﺴﺒﺖ ﺑﻪ اﻃـﺮاف آن ﻛﻤﺘـﺮ ﺑﺎﺷـﺪ (هوگنبوم، 1987). درواقع در ﺷـﺮاﻳﻂ ﺗـﻨﺶ ﺧﺸﻜﻲ ﭘﺘﺎﻧﺴﻴﻞ آب ﻣﺤﻴﻂ ﮔﻴﺎه منفیتر از ﺷﺮاﻳﻂ ﻃﺒﻴﻌﻲ ﺑﻮده و ﺟﺬب آب ﺗﻮﺳﻂ ﮔﻴﺎه ﺑﺎ ﻣﺸﻜﻞ ﻣﻮاﺟﻪ میشود. ﺗﺤﻘﻴﻘﺎت ﺑﺴﻴﺎري در ﻣﻮرد ﺗﺄﺛﻴﺮ ﻛﻤﺒﻮد آب ﺑﺮ رﺷﺪ و ﻧﻤﻮ ﮔﻴﺎﻫﺎن انجامشده اﺳـﺖ اﻳـﻦ ﺗﺤﻘﻴﻘـﺎت ﺣـﺎﻛﻲ از آن اﺳﺖ ﻛﻪ ﻛﺎﻫﺶ رﺷﺪ ﺑﻪ دﻻﻳﻞ ﻣﺨﺘﻠﻔﻲ ﺣﺎدث میشود. وﻗﺘﻲ ﮔﻴﺎﻫﺎن ﺑﻪ آب ﻛﺎﻓﻲ دﺳﺘﺮﺳﻲ ﻧﺪاﺷﺘﻪ ﺑﺎﺷﻨﺪ، ﻣﻘﺪار مواد ﺑﺎزدارﻧﺪه رﺷﺪ ازجمله آﺑﺴﻴﺰﻳﻚ اﺳﻴﺪ، در ﮔﻴﺎه اﻓـﺰاﻳﺶ مییابد. از ﻃﺮﻓـﻲ ﺑﺮﺧـﻲ از ﻣﺤﻘﻘـﺎن ﻛـﺎﻫﺶ ﻣﻘـﺪار هورمونهای ﻣﺤﺮك رﺷﺪ ﻣﺎﻧﻨﺪ اکسینها، جیبرلینها ﺳﻴﺘﻮﻛﻴﻨﻴﻦﻫﺎ در ﮔﻴﺎه را براثر ﻛﻤﺒﻮد آب ﮔـﺰارش کردهاند (فهیمی،1996).درﮔﻴﺎﻫﺎن ﻧﺨﺴﺘﻴﻦ آﺛﺎر ﻛﻤﺒﻮد آب بهصورت ﺑﺴﺘﻪ ﺷﺪن روزنهها ﺑﺮوز میکند. ازآنجاییکه ﺑـﺮاي اﻧﺠـﺎم ﻋﻤـﻞ ﻓﺘﻮﺳﻨﺘﺰ ﺗﺒﺎدﻻت ﮔﺎزي ﺿﺮوري اﺳﺖ، ﺑﻨﺎﺑﺮاﻳﻦ، در اﺛﺮ ﻛﻤﺒﻮد آب و ﺑﺴﺘﻪ ﺷﺪن روزنهها ﺗﺒﺎدﻻت ﮔﺎزي کاهشیافته و درنتیجه کربن مونواکسید ﻛﻤﺘﺮي در دﺳﺘﺮس ﮔﻴﺎه ﻗﺮار میگیرد و شدت ﻓﺘﻮﺳﻨﺘﺰ ﻛﺎﻫﺶ مییابد (مارتین و رزای،1992). ﺗﻨﺶ ﺧﺸﻜﻲ از ﻃﺮﻳﻖ ﻋﻮاﻣﻞ ﻏﻴﺮ روزنهای ﻧﻴﺰ ﺑﺮ ﺷﺪت ﻓﺘﻮﺳﻨﺘﺰ تاثیر میگذارد. ﺑﻄﻮرﻳﻜﻪ واکنشهای ﺑﻴﻮﺷﻴﻤﻴﺎﻳﻲ ﻓﺘﻮﺳﻨﺘﺰ همچنین دﺳﺘﮕﺎه ﻓﺘﻮﺳﻨﺘﺰي در اثر ﻛﻤﺒﻮد آب آسیبدیده و درنتیجه ﺷﺪت ﻓﺘﻮﺳﻨﺘﺰ ﻛﺎﻫﺶ مییابد. ﻋﻼوه ﺑـﺮ اﻳـﻦ در ﺷﺮاﻳﻂ ﺗﻨﺶ ﺧﺸﻜﻲ ﺳﻄﺢ ﺑﺮگ ﻧﻴﺰ کاهشیافته و اﻳﻦ امر نیز ﺑﺎﻋﺚ ﻛﺎﻫﺶ ﻓﺘﻮﺳﻨﺘﺰ ﺧـﺎﻟﺺ میشود (استراک، 1981). تنش خشکی با کاهش سطح برگ، انسداد روزنهها، کاهش فعالیتهای پروتوپلاسمی و تثبیت گازکربنیک، کاهش سنتز پروتئین و کلروفیل سبب تقلیل فرایند فتوسنتز میگردد (امام، 1383). جانسون و هرتز ثابت کردهاند که پتانسیل آب برگ میزان فتوسنتز را مستقیماً تحت تاثیر قرار میدهد. با افزایش تنش آب فتوسنتز تا نقطه فتوسنتز کاهش مییابد و بهطور مستقیم بر فرایندهای بیوشیمیایی مربوط به فتوسنتز اثر گذاشته و بهطور غیرمستقیم ورود گازکربنیک به داخل روزنهها را که به علت تنش آب مسدود باشند را کاهش میدهد (فائو، 2005). شرایط تنش شدید تنفس، جذب گازکربنیک، انتقال مواد فتوسنتزی و انتقال مواد خام در آوندهای چوبی به حد بسیار کم نزول کرده و این در حالی است که فعالیت آنزیمهای هیدرولیز کننده افزایش مییابد، کاهش مواد فتوسنتزی موجب اشباع برگها از این مواد میگردد. درنهایت کاهش فتوسنتز را در پی خواهد داشت (کوچکی،1365). بویر با تشریح فیزیولوژیکی غلات تحت تنش اظهار داشته تنش رطوبتی موجب کاهش فتوسنتز، پیری زودرس و ریزش برگهای پایین گیاه میگردد (بویر، 1995). در شرايط تنش كاهش ماده خشك میتواند به دليل فشار آماس سلول ناشي از كاهش سطح برگ گياه همچنين كاهش نرخ فتوسنتزی به دليل محدودیتهای بيوشيميايي ناشي از كمبود آب از قبيل كاهش رنگدانههای فتوسنتزی بهخصوص كلروفيل ها باشد (لاولر،2002). 1-2-5 تنش خشکی و اختلال در فعالیت گیاه تغييرات فيزيولوژيکی که در اثر کمبود آب در گياه ايجاد میشود، صدمات اکسيداتيو نيز از عوامل مهم محدودکننده رشد و توليدات گياهی هستند که در اثر عدم وجود شرايط مناسب ايجاد میشود. بيشترين صدمه در گياهان صدمه اکسيداتيو در سطح سلول است (آلن،1995). آلن گزارش کرده است که بيشترين خسارت در گياهان که از طريق تنشهای مختلف اعمال میشود، در ارتباط با خسارت اکسيداتيو در سطوح مختلف سلولی است. ارگانیزمهای فتوسنتز کننده هوازی در طول دوره زندگی در معرض ميزان متفاوتی از سمومی هستند که اين سمیت در گياهان عالی از کمبود آب حادتر است (سایرام وهمکاران، 2001). نتايج مطالعات نشان میدهد در شرايط تنش وحتی تحت غلظتهای بالای CO2 محيطی، بازهم فتوسنتز کاهش مییابد که بیانکننده اين امر است که دستگاه فتوسنتز صرفنظر از بسته شدن روزنهها، تحت تاثیر قرارگرفته است. در شرايط تنش کمبود آب روزنهها در گياه بسته میشوند و متعاقب آن غلظت CO2 در بافت مزوفيل کاهش مییابد و به دنبال اين وضعيت واکنشهای تاريکی فتوسنتز مختل شده و محصولات حاصل از واکنشهای روشنایی که شامل ATP، NADPH است، مصرف نمیشود. در چنين شرايطی به دليل عدم اکسيد شدن مولکول NADPH، مصرف NADP+ جهت دريافت الکترون کاهش مییابد، بنابراين مولکول اکسيژن در مسير زنجيره انتقال الکترون بهعنوان پذيرنده جانشين الکترون عمل میکند و منجر به شکلگیری راديکال سوپر اکسيد (O2-)، پر اکسيد هيدروژن (H2O2) و راديکال هیدروکسید (OH-) میگردد (سایرام و همکاران،2000؛تارکان،2005). فعاليت گونههای فعال اکسيژن[3] (ROS) ممكن است سبب بروز صدماتی همچون اكسيد شدن ليپيدها، (درنتیجه منجر به تغيير ساختار غشاء و درنتیجه ازهمپاشیدگی يكپارچگي آن میشود)، تغيير ساختمان پروتئینها و اكسيد شدن گروههای سولفیدریل (-SH)، غیرفعال شدن آنزیمها، بیرنگ شدن و یا از بين رفتن رنگدانه هادی مانند کلروفيل و ساير ترکيبات رنگیزهای و همچنین حمله مداوم به مولکولهای آلي مثل DNA و درنتیجه اختلال در رشتههای DNA گردد(حبیبی و همکاران، 2004؛ موهنتی، 2003؛ میتلر و همکاران، 2002).
1-2-6 مکانیسمهای مقاومت به خشکی در گیاهان گیاهان دارای مکانیزمهای مختلفی برای مواجه با تنش میباشند. ﻓﺮار از ﺧﺸﻜﻲ یکی از مکانیم هاست ﻛﻪ ﮔﻴﺎه ﺗﻼش میکند ﺑﺎ رﺷﺪ ﺳﺮﻳﻊ و کوتاهمدت ﺧﻮد ﻗﺒﻞ از ﺗﺨﻠﻴﻪ رﻃﻮﺑﺖ ﺧﺎك ﺑﺬرﺗﻮﻟﻴﺪ ﻛﻨﺪ. ﺗﺤﻤﻞ ﺑﻪ ﺧﺸﻜﻲ مکانیزم دیگری است که ﺗﻮﺳﻂ ﺗﻨﻈﻴﻢ اﺳﻤﺰي، اﻓﺰاﻳﺶ ﻗﺎﺑﻠﻴﺖ ارﺗﺠﺎع، ﻛﺎﻫﺶ اﻧﺪازه ﺳﻠﻮل و آب ﻛﺸﻴﺪﮔﻲ از ﻃﺮﻳﻖ ﺗﺤﻤﻞ ﭘﺮوﺗﻮﭘﻼﺳﻤﻲ و ﺗﻐﻴﻴﺮ در ﺧﺼﻮﺻﻴﺎت دﻳﻮاره ﺳﻠﻮﻟﻲ از روشهای ﺗﺤﻤﻞ ﮔﻴﺎه به شمارمی رود. (نیلسون و ارکوت، 1996؛ کوچکی و همکاران، 1372) ﺗﻨﻈﻴﻢ اﺳﻤﺰي با ﻛﺎﻫﺶ ﭘﺘﺎﻧﺴﻴﻞ اﺳﻤﺰي ﺑﻪ ﻋﻠﺖ ﺗﺠﻤﻊ ﻣﻮاد ﺣﻞ ﺷﻮﻧﺪه در ﭘﺎﺳﺦ ﺑﻪ ﻛﻤﺒﻮد آب ایجاد میشود. ﭘﺘﺎﻧﺴﻴﻞ اﺳﻤﺰي در داﺧﻞ ﺳﻠﻮل ﺑﺎ اﺿﺎﻓﻪ ﻳﺎ ﺣﺬف ﻛﺮدن ﻣﻮاد ﺣﻞ ﺷﻮﻧﺪه از ﺳﻠﻮل انجام میشود ﺗﺎ زﻣﺎﻧﻲ ﻛﻪ ﭘﺘﺎﻧﺴﻴﻞ اﺳﻤﺰي درون ﺳﻠﻮل ﺑﺎ ﭘﺘﺎﻧﺴﻴﻞ ﻣﺤﻴﻂ اﻃﺮاف ﺳﻠﻮل تقریبا ﺑﺮاﺑﺮ ﺷﻮد و اﻳﻦ ﻣﻮرد اﻏﻠﺐ ﺑﺮاي ﮔﻴﺎﻫﺎن ﭘﺴﺖ ﺑﻜﺎر میرود. ﭘﺘﺎﻧﺴﻴﻞ آب ﻛﻞ میتواند در ﻫﻨﮕﺎم بروز ﺧﺸﻜﻲ ﺗﻮﺳﻂ تنظیم اﺳﻤﺰي ﺑﺎ ﻛﺎﻫﺶ ﻣﻴﺰان پتانسیل آب ﺳﻠﻮل و ﻳﺎ افزایش مواد ﺣﻞ ﺷﻮﻧﺪه و با دﺧﺎﻟﺖ ﻗﻨﺪﻫﺎ ﺗﻨﻈﻴﻢ ﮔﺮدد و ﺑﺴﺘﻪ ﺑﻪ ﮔﻮﻧﻪ ﮔﻴﺎﻫﻲ، اسمولیتها اﺻﻠﻲ و ﻛﻠﻴﺪي در ﺗﻨﻈﻴﻢ اﺳﻤﺰي ﻣﺘﻔﺎوت اﺳﺖ. ﺑﺮاي ﻣﻮاد ﺗﺠﻤﻊ ﻳﺎﻓﺘﻪ اﺛﺮات ﻣﺨﺘﻠﻔﻲ گزارششده اﺳﺖ. اﻳﻦ ﻣﻮاد میتوانند ﺑﺎ اﻓﺰاﻳﺶ ﻧﮕﻬﺪاري آب و ﺣﻔﻆ ﭘﺘﺎﻧﺴﻴﻞ تورگر ﺳﺒﺐ ﺗﻨﻈﻴﻢ اﺳﻤﺰي ﺷﻮﻧﺪ. در ﺗﻌﺪادي از واکنشهای ﺑﻴﻮﺷﻴﻤﻴﺎﻳﻲ ﺟﺎﻧﺸﻴﻦ آبگردند و ﻫﻤﭽﻨﻴﻦ ﺑﺎ ﻟﻴﭙﻴﺪﻫﺎ ﭘﻴﻮﻧﺪ داده و از ﺗﺨﺮﻳﺐ ﻏﺸﺎء جلوگیری نمایند و ﻳﺎ از ﺗﻔﻜﻴﻚ کمپلکسهای ﭘﺮوﺗﺌﻴﻨﻲ و ﻳﺎ غیرفعال ﺷﺪه آنزیمها ﺟﻠﻮﮔﻴﺮي ﻛﻨﻨﺪ (زانگ و همکاران،1996). در ﺑﺮاﺑﺮ ﺗﻨﺶ ﺧﺸﻜﻲ واﻛﻨﺶ ﻋﻤﻮﻣﻲ ﺗﺠﻤﻊ ﻣﻮاد ﺣﻞ ﺷﻮﻧﺪه ﺳﺎزﮔﺎر ﺻﻮرت میگیرد ﻛﻪ ﺑﺴﻴﺎري از آنها اﺳﻤﻮﻟﻴﺘﻬﺎ ﻫﺴﺘﻨﺪ ﻛﻪ ﺑﺎﻋﺚ ﺗﻨﻈﻴﻢ اﺳﻤﺰي میشود. ﺗﺠﻤﻊ و ﻧﻮع ﻣﺎده ﺣﻞ ﺷﻮﻧﺪه در ﺑﺮاﺑﺮ ﺗﻨﺶ ﺑﻪ ﻣﻨﻈﻮر ﺳﺎزﮔﺎري در گونههای ﻣﺨﺘﻠﻒ ﻣﺘﻔﺎوت اﺳﺖ ﻛﻪ ﺷﺎﻣﻞ آﻣﻴﻨﻮاﺳﻴﺪﻫﺎ (پرولین), ﻗﻨﺪﻫﺎ (ساکارز، فروکتان) ﭘﻠﻲ مرها (مانیتور، سوربیتول) اسیدآمینه (ﮔﻼﻳﺴﻴﻦ، ﺑﺘﺎﺋﻴﻦ) یونها (ﭘﺘﺎﺳﻴﻢ)، اﺳﻴﺪﻫﺎي آﻟﻲ (مالت و ﻧﻴﺘﺮات) میباشند؛ مثل ﺗﺠﻤﻊ ﮔﻼﻳﺴﻴﻦ ﺑﺘﺎﺋﻴﻦ در ﺑﺮاﺑﺮ ﺗﻨﺶ اﺳﻤﺰي ﺧﺼﻮﺻﻴﺖ ﺑﺎرز در ﺧﺎﻧﻮاده Plumbaginaceae است درحالیکه در ﺗﻨﺒﺎﻛﻮ و ﺑﺮﻧﺞ ﺳﻴﺮ ﺑﻴﻮﺳﻨﺘﺰي ﮔﻼﻳﺴﻴﻦ بتائین دیده ﻧﺸﺪه اﺳﺖ (صفر نژاد، 1996؛رونتین همکاران، 2002). ﻛﻤﺒﻮد آب ﺳﺒﺐ ﺑﻴﺎن ژنهای ﺧﺎص میشود و ﺑﻴﺎن اﻳﻦ ژنها ﺑﺎ پاسخهای ﺳﺎزﮔﺎري ﮔﻴﺎﻫﺎن ﺗﺤﺖ تنش در ارﺗﺒﺎط اﺳﺖ (شین واری،2000). ﺗﻨﺶ آب ﺑﻪ ﺳﺒﺐ ﺗﻐﻴﻴﺮات ﺑﻴﺎن ژنها در ﮔﻴﺎﻫﺎن بهرغم ﻛﺎﻫﺶ ﺳﻨﺘﺰ ﭘﺮوﺗﺌﻴﻦ ﻛﻞ ﺑﺎﻋﺚ ﺗﻮﻟﻴﺪ پروتئینهای ویژهای میگردد ﻛﻪ سلولها و بهطورکلی ﮔﻴﺎه را در برابر تنش ﻣﺤﺎﻓﻈﺖ میکنند (صفرنژاد،1996). اﻳﻦ پروتئینها ﺷﺎﻣﻞ چاپرونها، LEAs (Late embryogenesis, abundant) اﺳﻤﻮﺗﻴﻦ و پروتئینهای ﺿﺪ ﻳﺨﺰدﮔﻲ، پروتئینهای متصل به mRNA و آنزیمهای ﻛﻠﻴﺪي ﺑﺮاي بیوسنتزی اﺳﻤﻮﻟﻴﺘﻬﺎ، پروتئینهای ﻛﺎﻧﺎل آب، ﻧﺎﻗﻠﻴﻦ پرولین و ﻗﻨﺪﻫﺎ و آنزیمهای سمزدا میباشند. پروتئینهای LEA،پروتئینهای ﻣﺘﺼﻞ ﺑﻪ mRNA و چاپرونها در ﻣﺤﺎﻓﻈﺖ ماکرو مولکولهایی ﻫﻤﭽﻮن آنزیمها، ﻟﻴﭙﻴﺪﻫﺎ و mRNA در ﺑﺮاﺑﺮ دﻫﻴﺪراﺳﻴﻮن دﺧﺎﻟﺖ دارﻧﺪ.پرولین، ﮔﻼﻳﺴﻴﻦ ﺑﺘﺎﺋﻴﻦ و ﻗﻨﺪﻫﺎ بهعنوان اسمولیتها ﻋﻤﻞ ﻛﺮده و از سلولها در برابر دهیدراسیون ﻣﺤﺎﻓﻈﺖ مینماید. پروتئینهای ﻛﺎﻧﺎل آب، ﻧﺎﻗﻠﻴﻦ ﻗﻨﺪﻫﺎ و ﻧﺎﻗﻠﻴﻦ پرولین در اﻧﺘﻘﺎل آب، ﻗﻨﺪﻫﺎ و پرولین ﺑﺎ ﻋﺒﻮر از ﻋﺮض ﻏﺸﺎﻫﺎي ﭘﻼﺳﻤﺎﻳﻲ و ﺗﻮﻧﻮﭘﻼﺳﺖ در ﺗﻨﻈﻴﻢ ﻓﺸﺎر اﺳﻤﺰي ﺗﺤﺖ ﺷﺮاﻳﻂ ﺗﻨﺶ ﻋﻤﻞ میکنند. آنزیمهای سمزدا شامل سوپر اکسيد دیسموتاز (SOD)، کاتالاز (CAT)، آسکوربات پراکسیداز (APX) و گلوتاتيون ردوکتاز (GR) میباشند در ﻣﺤﺎﻓﻈﺖ ﺳﻠﻮل در ﺑﺮاﺑﺮ اﻛﺴﻴﮋن ﻓﻌﺎل ﻋﻤﻞ میکند (یاماگیچی،2002). ﺗﻮﻟﻴﺪ پروتئینهای دﻳﮕﺮ ﻧﻴﺰ در ﺑﺮاﺑﺮ ﺗﻨﺶ آب ﻫﻤﭽﻮن (responsive to ABA) RABsdehydrins و پروتئینهای ذﺧﻴﺮه روﻳﺸﻲ در ﮔﻴﺎﻫﺎن ﻣﺨﺘﻠﻒ گزارششدهاند (Artlip and Funkhouser,1995). 1-2-3 آنزیمها آنزیمها کاتالیزورهای حیاتی و تمام پپتیدی یا پروتئینی هستند که توسط سلول زنده سنتز شده و عمل کاتالیز واکنشهای بیوشیمیایی را در داخل یا خارج سلول بر عهدهدارند. آنزیمها اختصاصیترین پروتئینهایی هستند که میتوان فعالیت آنها را بهصورت کاتالیز واکنشهای بیوشیمیایی نشان داد. این ترکیبات دارای ۲ خاصیت مهم هستند: 1- کاتالیز کردن واکنشهای بیوشیمیایی و افزایش سرعت این واکنشها. 2- عمل کردن اختصاصی بر روی پیش ماده، آنزیمها چنان اختصاصی عمل میکنند که میتوانند موادی را که دارای گردش نوری متفاوت هستند از یکدیگر بازشناخته و فقط بر روی ۱ نوع ایزومر نوری عمل کنند. در ساختمان آنزیم محلی مخصوص، که به آن جایگاه فعال میگویند وجود دارد. این جایگاه و همچنین شکل و گروههای شیمیایی ساختار آنزیم موجب میگردند که یک آنزیم بهطور اختصاصی بر روی یک سوبسترا عمل کند. ازنظر ساختمانی آنزیمها را به دودسته تقسیم میکنند. تعدادی از آنزیمها فعالیتشان تنها به ساختمان پروتئینی بستگی دارد، درحالیکه برخی دیگر از آنزیمها برای انجام فعالیتهای کاتالیزوری خود به ترکیبات فعالکننده غیر پروتئینی (کوفاکتور) نیاز دارند. کوفاکتورها نیز بر دو نوع میباشند: 1- کوفاکتورهایی که تنها شامل یک یون فلزی مانند Zn++، Mg++، Mn++، Cu++، Fe++، Na+ و یا k+میباشند. اینگونه کوفاکتورها به دو صورت عمل میکنند. الف: کوفاکتور نقش یک واسطه را برای پیوند مولکولهای آنزیم با سوبسترا بر عهدهدارند. ب: برخی از کوفاکتورها خود در عمل کاتالیز شرکت مینمایند. از این جمله میتوان فلز آهن را در آنزیم کاتالاز مثال زد. 2- کوفاکتورهایی که از یک مولکول ترکیب آلی تشکیل یافتهاند، اینگونه کوفاکتورها را کو آنزیم مینامند. کوفاکتورها اغلب با پیوند ضعیف و کم انرژی به قسمت پروتئینی آنزیم اتصال یافته است، ولی در بعضی موارد مشاهده میشود که این اتصال توسط پیوندهای کووالان انجامگرفته است. اینگونه کوآنزیمها را گروه پروستتیک مینامند. بیشتر کوآنزیمها از مشتقات ویتامینهای محلول در آب میباشند. کوآنزیمها عموما در مقابل حرارت مقاوم میباشند، درحالیکه قسمت پروتئینی (آپوآنزیم) در برابر حرارت ناپایدار و آسیبپذیر است.
1-2-3-1 اصول کلی واکنشهای آنزیمی الف: در واکنشهای بیوشیمیایی که توسط آنزیم کاتالیز می گردند، معمول سه عامل شرکت دارند: 1- سوبسترا یا جسمی که تحت تاثیرداشته آنزیم قرار میگیرد. 2- آنزیم یا ترکیبی که از آنزیم و کوفاکتور که بر روی سوبسترا اثر نموده و واکنش را کاتالیز میکند. 3- محصول که در جریان واکنش تولید میشود. ب: آنزیم در طی واکنش به مصرف نرسیده و بدون هیچگونه تغییری در خاتمه واکنش آزاد میگردد. 1-2-3-2 سوپراکساید دیسموتاز سوپراکسایددیسموتاز ها خط اول دفاعی گیاهان علیه رادیکالهای آزاد اکسیژن در درون سلول میباشند (افشر، 2002). سوپراکسایددیسموتاز آنزیمی متعلق به خانواده متالوآنزیمها است که رادیکالهای سوپراکساید را به پراکسیدهیدروژن و مولکول اکسیژن تبدیل میکند. این واکنش توسط کاتالاز و آسکوربات پراکسیداز ادامه مییابد یعنی آنها پراکسیدهیدروژن را به آب و اکسیژن تجزیه مینمایند و بدین ترتیب از تجمع رادیکالهای سوپراکساید که برای سلولها بسیار مخرب هستند، جلوگیری مینمایند (بیسر و همکاران،1991، بلوخینا و همکاران، 2003). سوپراکساید در هرجایی که چرخه انتقال الکترون وجود داشته باشد تولید میشود، بنابراین برانگیختگی اکسیژن میتواند در هر بخشی از سلول ازجمله میتوکندری، کلروپلاست، گلیاکسیزوم، پراکسی زوم، آپوپلاست و سیتوسل رخ دهد، درنتیجه نباید از وجود سوپراکساید دیسموتاز در تمام این اندامکها شگفتزده شد (افشر، 2002). بر اساس نتایج مطالعات آسادا و تاکاهاشی (1987)، غشاهای فسفولیپیدی توانایی تراوش مولکولهای O2- را ندارند، بنابراین وجود سوپراکساید دیسموتاز در محلی که مولکولهای -O2 تولید میشوند بسیار حیاتی است. افزایش بیشتر مقادیر آنزیم سوپراکساید دیسموتاز در واریتههای متحمل نخود، پنبه، گوجهفرنگی و ... در مقایسه با واریتههای حساس، میتواند دلیلی برای نقش مهم این آنزیم برای مقابله با تنش خشکی باشد (میتووا وهمکاران، 2002). برای توجیه این مسئله، به نظر میرسد که بالاتر ب انتشار : ۱۱ آبان ۱۳۹۶ برچسب های مهم پروژه خازن گذاري در شبكه هاي توزيع براي كاهش تلفات و بهبود ضريب توان
انتشار : ۱۱ آبان ۱۳۹۶ برچسب های مهم روش هاي ازدياد برداشت از مخازن نفتي
انتشار : ۱۱ آبان ۱۳۹۶ برچسب های مهم جزوه موازنه جرم و انرژی
انتشار : ۹ آبان ۱۳۹۶ برچسب های مهم تأثیر كاربرد کولتیواتور و چند کشتی همزمان بر خصوصیات زراعی و زیست توده ذرت در اصفهان
|














 اسمت را زیبا کن مخصوص تلگرام *****
اسمت را زیبا کن مخصوص تلگرام *****

 کسب درامد از اینترنت ماهیانه 8میلیون تومان *****
کسب درامد از اینترنت ماهیانه 8میلیون تومان *****



 ربات پیشرفته اینستاگرام *****
ربات پیشرفته اینستاگرام *****








